General behavioral responses (avoidance and aggregation)
Wilson and Regnier38 classified the alarm responses in ants as either “panic” or “aggressive”. As the specific alarm signaling in termites (comparable to panic in ants) is a subtle behavior71,72 out of the scope of this work, general alarm is accompanied by a dramatic change in the group behavior. The general alarm typically involves many individuals disturbed at foraging sites, or present in a part of the nest that has been locally damaged27,36,59,60,63,64,65,66,67,68,69. The alerting termites search for quiet termites, touch them with their antennae, and perform tremulations to alert them36. The alarm responses usually result in a high soldier recruitment activity at disturbance locations, where soldiers displayed defensive postures, often combined with the release of defensive secretions produced by the labial glands during the opening/closing of mandibles26,27. Locomotion activity increased in many cases, especially in Reticulitermes, and disturbed workers usually displayed higher locomotion activity (evasion) than soldiers (defensive confrontation; Fig. 1 and Tables S1 and S2).
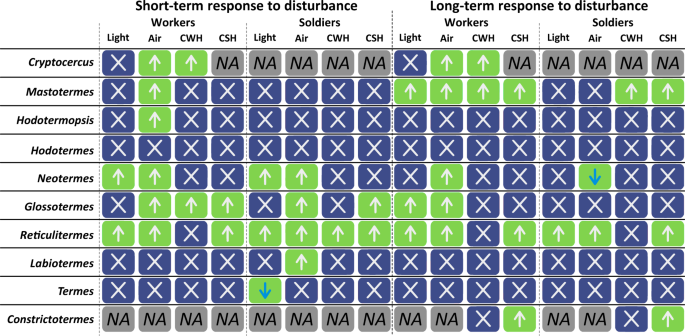
Fig. 1: Short-term (left) or long-term (right) responses in the locomotion speed in groups of nymphs of the wood roach Cryptocercus, or of workers and soldiers of different termite genera after exposure to different stimuli (light flash (light), air current (air), crushed worker head (CWH), and crushed soldier head (CSH)). Locomotion speed was recorded separately for workers (n = 2) and soldiers (n = 2). The green fields indicate significantly different locomotion speed after stimulation, upward arrows mean the speed increased, downward arrows mean the speed decreased, the blue fields indicate no significant difference, NA means data not available. Full size image
Workers attempted escaping from the source of disturbance by moving away rapidly, while soldiers often searched for the source of disturbance and aggregated around it, resulting in a slower-motion patrolling behavior usually combined with scanning of the space with wide-spread antennae and mandibles ready to be triggered (when present). Soldiers of the majority of the studied species with biting-type mandibles started opening mandibles after direct disturbance because of two reasons: (i) open mandibles are prepared to bite once the opponent reaches the antennae, (ii) repeated openings of mandibles stimulate the release of defensive chemicals from cephalic glands as the mandibular muscles squeeze the liquids out of reservoirs. The defensive secretion is usually delivered to the opponent together with the bite34,47,54,73. The response to disturbance also included higher production of vibroacoustic alarm by disturbed individuals that warns nestmates (Table S3). Aside from such common behavioral responses, more specific actions were repeatedly observed. Workers and soldiers of Hodotermopsis frequently showed a type of drumming that substantially differs from that of all other termites—vigorous oscillatory movements against the lid of the Petri dish (not against the ground as in all other cases). While the increased locomotion was mainly observed in response to direct disturbance, there were a few exceptions, as in Neotermes and Termes, in which soldiers significantly decreased their locomotion speed, although such change was observed in Neotermes in long-term response only, implying that the patrolling behavior followed the active search for the source of disturbance. Finally, Hodotermes was a remarkable outlier in its general behavioral response to disturbances, as both castes stopped all movements after the disturbance for a short time (freezing behavior), but resumed their previous activity within 1–2 seconds. Thus, based on our experiments and repeated field observations, it is likely that the escaping behavior (higher locomotion speed) of workers and the aggregation of soldiers towards a disturbance is a basal trait to all extant termites that was secondarily lost only once, in Hodotermes (Fig. 1 and Table S1).
Alarm pheromones
Alarm pheromones in termites originate from soldiers’ defensive glands only: the labial glands in Mastotermes27 and the frontal gland in Neoisoptera (the derived group comprising Stylotermitidae, Rhinotermitidae, Serritermitidae, and Termitidae)36,59,63,64,65,67,68,69. Similar signals are widely used in some cockroaches, produced by the abdominal sternal or tergal glands (Eurycotis74; Therea75; Blaberus76). Benzoquinone, the alarm pheromone of Mastotermes, originates from the soldiers’ labial glands, and triggers a typical alarm behavior including caste-dependent change in locomotion speed and increased vibroacoustic signaling. All other alarm pheromones originate from the frontal gland, a termite-specific organ with no equivalent in other groups77. The frontal gland of soldiers is a saccular gland that opens to the exterior through the fontanelle in all Neoisoptera species we studied but Glossotermes, which has a blind-ended sac in the thorax and abdomen, whose contents can eventually be released through self-sacrifice via body rupture34,73.
Our study included three genera of Neoisoptera using alarm pheromones—Prorhinotermes, Reticulitermes, and Constrictotermes (Fig. 2 and Table S1), along with data from the literature. The active components of alarm pheromones are terpene hydrocarbons in all Neoisoptera34,41,68. Although Glossotermes increased the locomotion speed in response to a crushed soldier head, our chemical analyses did not reveal any alarm pheromone candidate (Fig. S1, Table S4), in line with the lack of a frontal gland reservoir in its head73. However, more species revealed responses to crushed heads devoid of defensive glands or conspicuous volatiles (see Fig. 1 and Table S1), possibly because colony members can perceive the smell of dead or wounded termites78,79,80,81. The two soil-feeding species we studied (Labiotermes and Termes) lack alarm pheromones, unlike wood-feeders that used alarm pheromones. In addition to field observations, it suggests that alarm pheromones are not used by the soil-feeding groups, representing altogether 60% of termite species diversity70. According to recent termite phylogenies82,83, alarm pheromones evolved at least twice; in the most basal extant termite clade, Mastotermitidae, and then in Neoisoptera, in which the lack of observations precludes our distinguishing between a single origin followed by multiple losses or multiple origins (Fig. 2). Our investigation suggested that the species’ life-type and related traits strongly correlated with the presence of alarm pheromones (Fig. S2), as species living either in small colonies (Archotermopsidae, Kalotermitidae70,84), or those living in open arid environments (Hodotermitidae70,84), lack this communication channel.
Fig. 2: General comparison of the evolutionary history of vibroacoustic and chemical alarm distribution among termites and the wood roach Cryptocercus. Dots are yellow when the alarm mode is present, white when absent, and gray when unknown. The family name colors represent ranks for MCA analysis. Full size image
Vibroacoustic signaling
Termites generally responded to disturbance by violent shaking and drumming, sometimes accompanied by sounds audible to the observer (Movie S1). The beats were arranged into bursts of low frequency (under 15 Hz) in the case of low tremulation, or high frequency (above 15 Hz) high tremulation, drumming or head-banging. The vibroacoustic signature was specific to a given genus (Figs. 3, 4, and S3), a feature not previously recognized.
Fig. 3: Scheme of the main features of the vibroacoustic alarm communication linked to phylogeny (n = 50 for each species and castes). In contrary to Fig. 2, we included only species for which vibroacoustic communication is known in sufficient details. Full size image
Fig. 4: Frequency (in Hz) of low tremulations (yellow background) and high (green background) tremulations (n = 50 for each species and castes) across studied species. The nymphs of the wood roach Cryptocercus and termite workers in black, and drumming by termite soldiers (in white). On each box, the central mark is the median, the edges of the box are the 25th and 75th percentiles, the whiskers extend to the most extreme datapoints the algorithm considers to be not outliers, and the outliers are plotted individually. Cryptocercus and Neotermes produced low frequency signals only. Hodotermes is not included as it does not communicate via body vibrations. Full size image
While the species descending from early diverging lineages (including Cryptocercus85) revealed a rather monotonous pattern of beats arranged into singular bursts, the patterns became more diverse in Neoisoptera, such as in Glossotermes, and especially in Termitidae, which use a combination of several bursts into a single vibroacoustic event (Figs. 3 and S3). The tremulations were primarily used as short-range communication to alert naïve nestmates, and carried relatively low energy compared to drumming or head-banging. The tremulations, when processed and amplified, are audible as muffled noise, while drumming and head-banging sounds like a series of sharp hits (Movie S1). The occurrences of respective signal components are summarized in Table S1. Workers and soldiers within a species mostly share the same repertoire, although additional signals, such as head-banging, occur in soldiers only. The general trends show that the larger species vibrate at lower frequencies, and Cryptocercus and Neotermes, both high above the average termite size, lack the high-frequency signals completely. Hodotermopsis is unique among termites for hitting its head against the ceiling for drumming, not the floor, in both workers and soldiers. Head-banging was abundantly recorded in Glossotermes and Reticulitermes (Figs. 3, 4, and S3), but was only rarely observed in Labiotermes. All Termitidae displayed relatively conserved and complex patterns of vibroacoustic sequences combining abdomen drumming, and high and low tremulations into long sequences (Movie S1, Figs. 3, and S3). Notably, vibroacoustic signals were extremely well conserved within species, with little variation among subsequent beats, showing that alarm signals have been quite stable since Cryptocercus and termites diverged (Figs. 5 and S4), an event dating back to at least the Late Jurassic82,83. We may therefore assume that the pre-social ancestor of Cryptocercus and termites used alarm signals of comparable precision.
Fig. 5: Stability of breaks between two subsequent beats in low tremulations across the species (n = 50 for each species). The values are given as percentage difference (positive or negative) from the mean duration. On each box, the central mark is the median, the edges of the box are the 25th and 75th percentiles, the whiskers extend to the most extreme datapoints the algorithm considers to be not outliers, and the outliers are plotted individually. Full size image
Evolutionary trajectories of alarm signals in termites
Our ancestral-states reconstructions indicate a single origin of vibroacoustic alarm communication in the common ancestor of all termites and their sister group, the wood roach Cryptocercus (Fig. 2). Its loss in Hodotermes (and Anacanthotermes, D.S.D. and J.Š., field observation) is probably due to environmental conditions: nesting in soft sandy ground and randomly foraging in the open air to collect dry grass presumably prevents effective transmission of vibrations or odors. To our best knowledge, the loss of vibroacoustic alarms occurred exclusively in Hodotermitidae. Whether vibratory communication disappeared completely in this group, or whether it was partly retained in another social context such as nest defense, as it is common in other termites86,87,88,89, remains to be determined.
Even though vibroacoustic communication is shared by all colony members, the actual involvement of the different castes in alarm signaling has rarely been studied. Our data show that both castes mostly share identical part on the communication, except in Glossotermes, Reticulitermes, and Labiotermes, in which soldiers perform head-banging, a soldier-specific behavior. Apart of these differences, our data do not support increased involvement of soldiers in alarm signaling, in contradiction to observations made by Stuart90. We could observe increased diversity of alarm signals in derived termite taxa, evidenced mostly in vibroacoustic alarm sequences, since alarm pheromone data are made only in the presence/absence of this channel.
Multiple Correspondence Analysis (MCA) analysis suggested that several ecological characteristics have a strong influence upon vibroacoustic communication. The most prominent ecological characters are the hardness of the food and the nest material, which, along with phylogenetic position, are the most important features influencing vibroacoustic signaling (Figs. 6 and S5), probably because relatively hard substrates facilitate the transmission of such signals. In addition, we found a clear relationship between the frequency of oscillatory movements used in vibroacoustic communication and termite body size, since larger species always communicate at lower frequencies than smaller ones.
Fig. 6: Square correlation ratio between characters and axis in MCA. Both graphs are in fact the same, and the left one shows the most influential characteristics of vibro-acoustic features (in blue) to the MCA analysis and the right one shows the most important ecological characters (in green) that might have an effect on vibroacoustic traits. The larger the size of the letters, the more important the character. Full size image
Pairwise correlation analyses showed that vibroacoustic characters are strongly correlated between the soldier and worker castes. Moreover, it seems that the presence of tremulation correlates with drumming in both castes across all species, and that the presence of tremulation in the soldier caste is correlated with the size ratio between castes (Fig. S2). Termite taxa with proportionally large soldiers compared to workers, such as Hodotermopsis or Neotermes, primarily rely on soldier behavior for the spread of alarm, while termite taxa in which the soldier and worker castes have similar sizes rely equally on soldiers and workers to communicate. There is certainly a phylogenetic component to this as soldiers in more basal lineages tend to be larger, but it possibly also reflects the fact that heavier individuals spread vibroacoustic alarm more efficiently, rendering the spread of alarm communication by smaller individuals obsolete. Whether the specialization for alarm transmission of larger individuals holds or not among subcastes of species with polymorphic soldiers, such as some Macrotermitinae (Macrotermes, Acanthotermes, Ancistrotermes) or some Nasutitermitinae (Diversitermes, Trinervitermes), remains to be investigated.
An interesting correlation exists between the presence of chemical alarm and the colony life-type (Fig. S2). One-piece nesters living in hard wood (common among basal termites) do not utilize chemical alarms, probably because vibroacoustic alarm communication is sufficient in small colonies sheltered in sound wood84,91. These species usually defend against intruders at a few “bottleneck” entrance points in the gallery network92. Termite colonies contained within a small gallery system of a single piece of wood also use vibrations for purposes other than alarm communication. For example, Cryptotermes spp. may evaluate a looming resource shortage and perceive approaching competitors through substrate vibrations, which may result in the initiation of mass production of alates for a final dispersal flight30,31. Our data suggest that chemical alarms have only emerged in termite species that are able to use food resources outside their nest location, although not all central site-nesting termites appear to possess these. Species colonizing new food sources through underground foraging galleries are more likely to encounter enemies inside their galleries93, potentially increasing the selection pressure for the acquisition of an alarm pheromone. The use of volatile alarm pheromones may efficiently alert naïve individuals of a potential threat when they approach a disturbed area, while soft substrates may not favor vibroacoustic communication. We can speculate that alarm pheromones may persist in the air even after the death of the foraging termites, triggering local avoidance of the newcomers.
The identification of alarm pheromones is intricate, and therefore they have been identified for only a few species27,36,59,65,68,94. Here, we have not studied the chemical nature of alarm pheromones, but merely their presence/absence based on the soldier crushed heads, behavioral responses to them, and the composition of their volatiles. A maximum parsimony model supports independent evolutionary origins of chemical alarms in Mastotermitidae and Neoisoptera, followed by repeated losses in the latter lineage. Many termite and cockroach species produce defensive compounds, often irritating, that, presumably, can be co-opted as alarm pheromones, explaining the diversity of alarm pheromones and their glandular origins found among Blattodea under highly variable selective pressures27,34,74,75,76.