In vitro resistance of GAS to SXT requires high-level TMP resistance conferred by dfrF
To investigate the combinations of TMP and SMX resistance that confer SXT resistance, we determined the susceptibility for SMX, TMP and SXT for a global collection of GAS strains reported to have reduced susceptibility to SXT13,15 (Supplementary Table 1). The minimal inhibitory concentration (MIC) for each strain was determined using Epsilometer test (Etest) strips and Mueller-Hinton Fastidious agar (MHF; BioMerieux). While each strain was originally reported as being SXT-resistant using broth microdilution methodology13,15, only 5/22 were SXT-resistant using Etest methodology. This discrepancy is potentially explained by methodological differences between these assays, or differences in the composition of the growth medium used in each assay.
To identify genetic determinants associated with SXT resistance, we next compared MIC values for each antibiotic with known antibiotic resistance genes identified by whole genome sequencing (Supplementary Table 1). Supporting previous reports12,13, we identified TMP-resistance genes dfrF and dfrG in GAS isolates that exhibited TMP MIC values >32 μg/ml. dfrF and dfrG were variably encoded on horizontally acquired genetic elements (e.g. integrative conjugative elements), often with a number of additional antibiotic resistance genes. While several strains were found to have SMX MICs >100 μg/ml that were sufficient for SXT resistance, we were unable to identify any horizontally-acquired dihydropteroate synthase variants that would account for SMX resistance in these strains. Inspection of the sequence of the SMX target enzyme (FolP) in each strain revealed considerable sequence variation, yet there were no individual amino acid sequence variations that correlated with reduced SMX susceptibility (Supplementary Fig. 1). Structural16 and biochemical17 investigations have identified key residues in dihydropteroate synthase enzymes involved in SMX drug binding and resistance, which correspond to residues F25, S26, T59 and P61 in the GAS FolP sequence (Supplementary Fig. 1). Of these, only T59 showed any variation between the strains investigated, however the two amino acid variations at this residue did not correlate with the observed SMX MICs for strains that contained them (T59A, MIC range 48 to >1024 μg/ml; T59N, MIC 48 μg/ml). Thus, while we can not discount that combinations of FolP amino acid changes might have contributed to a reduced SMX susceptibility in some of these strains, the exact mechanism of resistance remained unclear.
dfrF confers much higher levels of TMP resistance than dfrG
Comparison of SXT susceptibility results for each strain in Supplementary Table 1 revealed that only dfrF-positive strains were SXT-resistant (MIC > 2 μg/ml). Surprisingly, both dfrG-positive strains (TB08 and TB10) were highly-resistant to TMP (MIC > 32 μg/ml) and SMX (MICs > 1024 μg/ml) yet remained SXT-sensitive. As TMP is a competitive inhibitor of substrate binding to dihydrofolate reductase enzymes, resistant enzyme variants have reduced affinity for TMP such that higher levels of antibiotic are required to inhibit enzyme activity18. As each dfrF- and dfrG-positive GAS strain had TMP MICs greater than the highest level resolved with the TMP Etest (32 μg/ml), we next compared levels of TMP susceptibility using a gradient plate assay (Supplementary Fig. 2A). Both dfrG-positive GAS strains were resistant to TMP concentrations between 50 and 200 μg/ml, while each of the dfrF-positive strains were resistant to more than 1000 μg/ml. Thus, the SXT-resistant phenotype of dfrF-positive GAS is likely attributed to their very high levels of TMP resistance, while the SXT-sensitive phenotype of dfrG-positive GAS is likely attributed to the lower levels of TMP resistance combined with highly synergistic activity of SMX and TMP19 (Supplementary Fig. 2B).
Forced evolution of dfrG-positive GAS strain TB08 leads to SXT resistance through upregulation of a ECF transporter substrate-binding component (thfT)
While dfrG-positive GAS isolates were sensitive to SXT, we reasoned that they might have the capacity to evolve SXT resistance with strong antibiotic selection. To investigate this, we examined the ability of a dfrG-positive/SMX-resistant/SXT-sensitive strain (TB08) to evolve SXT resistance following 20 rounds of serial passage on inhibitory levels of SXT. To do this, a single TB08 colony was spread over the surface of an agar plate containing a gradient of SXT (0–50 μg/ml) and cultured overnight. After each round of growth, colonies that grew at the highest concentration of SXT were used to seed subsequent rounds of evolution. An overview of this process is illustrated in Fig. 2a. After each round of selection, SXT susceptibility was monitored on SXT gradient plates (Supplementary Fig. 3) and by Etest (Fig. 2b). In one of three biological replicates, we observed a switch of TB08 to a SXT-resistant phenotype with the gradient plate assay (Supplementary Fig. 3), however a comparable SXT-resistant switch was not observed by Etest (Fig. 2b, blue circles).
Fig. 2: A DfrG-positive GAS strain can evolve SXT resistance due to upregulation of an ECF transporter S component gene (thfT). a Schematic of in vitro evolution procedure using SXT gradient plates. An overnight culture of TB08 on HBA was used to inoculate a SXT gradient plate (0–50 μg/ml) and incubated for 24 h at 37 °C. Growth was then harvested from the the highest antibiotic concentration (i.e. where growth started to be inhibited) and used to seed successive rounds of evolution. Created with BioRender.com. b MICs to SXT on MHF agar from bioMerieux (MHF-Bm; blue circles) and Oxoid (MHF-Ox; red squares) following in vitro evolution of GAS strain TB08. Results from three independent experiments are shown. Values are expressed as a ratio of the initial MIC as determined by Etest following each round of evolution. c Genomic context of thfT (purple arrow) as determined by tblastX pairwise blast which highlights evidence of horizontal gene transfer of thfT (purple) and shared origin with that in Streptococcus dysgalactiae subsp equisimilis. Percent identity is by shading as indicated to the right. d Relative expression of thfT in TB08 and TB08-2-14 measured by RT-qPCR. Values are normalised to average expression of TB08 ME samples (dashed line). ME, mid-exponential phase (OD 600 = 0.4–0.6); St, stationary phase (OD 600 = 1.2–1.4). Data from three biological replicates are presented as mean values ± SD. Differences assessed using a one-tailed, unpaired Student’s t-test. Source data for panels b and d are provided as a Source Data file. Full size image
In the in vitro evolution experiments above, the gradient plate (Supplementary Fig. 3) and Etest (Fig. 2b) assays used to monitor SXT susceptibility used different formulations of growth media. Gradient plates were prepared using Mueller-Hinton (MH) agar base (Oxoid) containing 2.5% lysed horse blood, while Etest assays were conducted on commercial Mueller-Hinton Fastidious agar plates (Mueller-Hinton agar containing 5% defibrinated horse blood and 20 μg/ml β-NAD from BioMerieux; MHF-Bm). To examine whether differences in media composition accounted for the differences in SXT MICs between the gradient plate and Etest assays, we next measured SXT MICs using Etest strips on MHF prepared using MH agar base from Oxoid (MHF-Ox; Fig. 2b, red squares). This analysis showed that the observed switch of TB08 to a SXT-resistant phenotype was dependent on the media composition, and only observed on MHF-Ox. Further examination of the TB08 parent and the evolved TB08 (from Experiment 2, round 14; strain TB08-2-14) revealed that TB08 and TB08-2-14 remained SXT-sensitive on MHF-Bm, while TB08-2-14 was resistant on MH agar from Oxoid containing different combinations of horse blood, lysed horse blood and β-NAD (Supplementary Fig. 4). These results demonstrate that the dfrG-positive/SMX-resistant isolate TB08 can switch to a SXT-resistant phenotype under strong selective pressure, and detection of this switch depends on the composition of the growth medium.
To identify the mechanism of evolved resistance in TB08-2-14, we performed whole genome sequencing on this strain and compared this sequence with the parent strain TB08. This comparison revealed that the SXT-resistant phenotype of TB08-2-14 was associated with a 5 bp sequence duplication in the upstream intergenic region of a putative folate transporter gene (herein named thfT). thfT encodes a protein with homology to FolT, the folate-specific substrate-binding component (S component) of an energy-coupling factor (ECF) transporter20,21 from Lacticaseibacillus casei22 (28% identity, 46% similarity) and Leuconostoc mesenteroides20 (23% identity, 45% similarity; Supplementary Fig. 5). Comparison with other SXT susceptible GAS genomes (e.g. MGAS5005, Fig. 2c), revealed thfT is inserted between murF and ddlA, two genes that comprise part of the core GAS genome and encode enzymes involved in peptidoglycan biosynthesis.
The 5 bp duplication in TB08-2-14 is in the intergenic region between thfT and the divergently transcribed gene ddlA. As this intergenic region likely contains the promoter region for thfT, we next examined whether the increased SXT resistance in TB08-2-14 was associated with upregulation of the thfT gene. RNA was prepared from each strain and thfT mRNA levels were quantified by qRT-PCR (Fig. 2d). Results from this experiment demonstrate that the 5 bp duplication in TB08-2-14 is associated with an increased level of thfT transcript in both mid-exponential (16.6-fold) and stationary (3.8-fold) phases of growth relative to TB08. This suggests that thfT could be an AMR gene that is up-regulated in TB08-2-14 due to a mutation in the thfT promoter region.
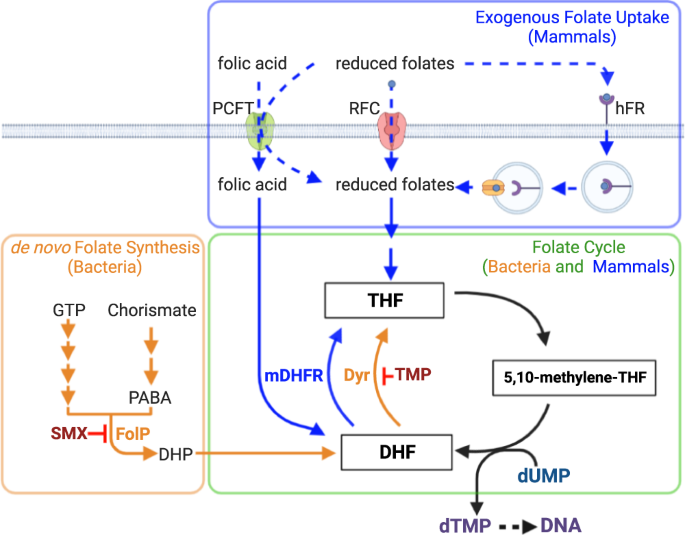
ThfT enables GAS to bypass the inhibitory effects of SXT through acquisition of extracellular tetrahydrofolate (THF)
The above results suggest that increased expression to thfT in TB08-2-14 is responsible for the increased SXT resistance of this strain relative to the parent strain TB08. Given the homology of ThfT to the substrate binding S component of an ECF folate transporter, we hypothesised that ThfT might expand the substrate profile of an endogenous ECF transporter to include end products of the folate biosynthesis pathway. This would enable thfT-positive GAS strains to bypass the inhibitory effects of SMX and/or TMP (Fig. 1). To examine this possibility, we first tested SXT MICs for TB08 and TB08-2-14 and a thfT-negative strain (M6JRS4) in the presence of different amounts of THF (Fig. 3a, b; Supplementary Fig. 6) and folic acid (Supplementary Fig. 7). While the thfT-negative strain M6JRS4 did not exhibit any change in SXT MIC in the presence of either compound, both TB08 and TB08-2-14 showed an increased SXT MIC in the presence of extracellular THF but not folic acid, with both strains being resistant to >32 μg/ml SXT at THF concentrations >50 ng/cm2 THF. The lack of resistance in the presence of extracellular folic acid likely reflects that folic acid is a very poor substrate for most bacterial dihydrofolate reductase enzymes23. A dose-response comparison by Etest showed that the amount of THF required to increase SXT resistance in the evolved strain TB08-2-14 (EC 50 = 3.37 ng/cm2; Fig. 3a) was lower than the amount required for the parent strain TB08 (EC 50 = 24.7 ng/cm2; Fig. 3a). Similar results were obtained by broth microdilution assay (TB08-2-14 EC 50 = 1.72 ng/ml, TB08 EC 50 = 7.60 ng/ml; Supplementary Fig. 6). These results are likely explained by the increased levels of ThfT in TB08-2-14, which may then outcompete other ECF S components for the same ECF transport module (Fig. 3c). This is supported by recent evidence that ECF S components can compete for the same ECF transport module, with high levels of substrate for one S component (e.g. pantothenate) able to reduce uptake of second substrate (e.g. folate) that is transported by an alternative S component24. Together, these data indicate that THF or related compounds are likely present in MH agar sourced from Oxoid at an intermediate concentration sufficient to support SXT resistance for TB08-2-14 but not for TB08 (Fig. 3a). In contrast, these compound(s) are absent or at much lower concentrations in MHF agar from BioMerieux (MHF-Bm) and are unable to support SXT resistance for either strain.
Fig. 3: ThfT confers SXT resistance in the presence of exogenous THF. a Dose-response curves for the impact of exogenous THF on SXT resistance of M6JRS4, TB08 and TB08-2-14. MICs are expressed as a percentage the maximum resolution of the Etest assay (32 μg/ml). Data from three biological replicates on MHF-Bm agar are presented as mean values ± SD. EC 50 values were calculated using Graphpad Prism software. b Representative Etest assays on MHF-Bm agar for data presented in a. c Schematic showing predicted effect of increased ThfT expression in TB08-2-14 on competition with other ECF S components for ECF transport modules and the effect on SXT MICs. Under conditions of low ThfT expression and low exogenous THF, competition between THF-loaded ThfT and S components loaded with other compounds limits THF import, resulting in low SXT MICs (top left). This competition is overcome by increased ThfT expression (top right) or increased extracellular THF, which results in high SXT MICs. Source data for panel a is provided as a Source Data file. Full size image
ThfT is a SMX resistance protein that confers very high levels of SMX resistance
Comparative genomics of 24 globally sourced GAS strains (Supplementary Table 1) revealed five strains that encoded thfT. To investigate whether all strains that encoded thfT were SXT resistant with exogenous THF, we next measured SXT MICs of each strain in the absence and presence of 50 ng/cm2 THF (Supplementary Table 2). Results showed that each of the thfT-positive strains that also encoded a TMP resistance gene (dfrG or dfrF) and were TMP resistant (>32 μg/ml) became highly-resistant to SXT in the presence of exogenous THF, while both of the strains that lacked a TMP resistance gene exhibited a modest (~4 to 5-fold) increase in SXT MIC values in the presence of THF.
Alignment of the predicted ThfT amino acid sequences identified two allelic variants that differed by a single amino acid (T71 and A71). Each of the strains with the T71 variant also encoded a TMP resistance gene, while both strains with the A71 variant lacked a TMP resistance gene and were TMP sensitive. To rule out any effects in affinity for THF in the A71 variant that might explain the different phenotype of the strains that encoded each variant, we examined SXT resistance across a range of THF concentrations (Supplementary Table 3). Addition of increasing extracellular THF (up to 400 ng/cm2) was unable to confer high-level SXT resistance for either of the A71 variant strains, suggesting that the lower level of SXT resistance in the A71 strains was not due to a reduced affinity for THF. Together, these results suggest that ThfT may be functioning as a SMX resistance protein, and that ThfT-mediated SXT resistance requires an additional mechanism of TMP resistance.
To further investigate whether ThfT conferred SXT resistance or SMX resistance, we expressed each ThfT variant in a laboratory GAS strain (M6JRS4) that lacked any TMP- or SMX-resistance mechanism, and measured the effect of exogenous THF on MIC values for SMX, TMP and SXT (Fig. 4a, Supplementary Table 4). Expression of either variant resulted in a very modest increase in SXT MIC in the presence of 50 ng/cm2 of exogenous THF, yet had no effect on MIC values obtained for TMP. However, expression of either variant resulted in high level SMX resistance (>1024 μg/ml) even in the absence of additional extracellular THF. This result was unexpected since addition of exogenous THF is required for SXT resistance in the parent strain (Fig. 3a, b). We rationalised that the lack of a requirement for additional THF for SMX resistance in this experiment may be explained by higher expression levels of thfT when expressed on a multicopy plasmid, which may then require lower levels of exogenous THF to bypass SMX inhibition of the folate synthesis pathway. The requirement of only small amounts of extracellular THF is likely explained by the fact that THF is recycled back to DHF during the production of thymidine monophosphate, such that only very low levels of THF should theoretically support DNA synthesis (Fig. 4b).
Fig. 4: ThfT is an SMX resistance protein that allows GAS strains to bypass the folate synthesis pathway in the presence of exogenous THF. a Heterologous expression of T71 and A71 variants of thfT in M6JRS4. The constructs with a constitutive P23 promoter are indicated with a blue arrow and thfT is indicated with a red arrow. SMX, TMP and SXT MICs were determined by Etest. Exogenous THF was supplied at a concentration of 50 ng/cm2. Results are median values of three independent experiments determined with Etest strips on MHF-Bm agar. Statistical analysis of groups is in Supplementary Table 4. b Model of ThfT mediated SMX-resistance. ThfT is able to acquire extracellular THF which then feeds directly into the folate cycle (blue box), which bypasses the inhibitory effect of SMX on folate synthesis (green box). As THF is recycled back to DHF during the folate cycle, only small amounts of exogenous THF are required to support growth in the presence of SMX. GTP, guanosine triphosphate; DHP, dihydropteroate; DHF, dihydrofolate; dUMP, deoxyuridine monophosphate; dTMP, deoxythymidine monophosphate. Created with BioRender.com. c Single gene complementation of thfT in the same genetic locus as in TB08 is sufficient for high-level SMX resistance in GAS strain NS5347. SMX, TMP and SXT MICs were determined by Etest. Exogenous THF was supplied at a concentration of 50 ng/cm2. Results are median values of three independent experiments determined with Etest strips on MHF-Bm agar. Statistical analysis of groups is in Supplementary Table 5. d Ability of THF to confer ThfT-mediated SMX resistance as measured by broth microdilution. Data from three biological replicates are presented as mean values ± SEM. Differences assessed using a two-tailed, unpaired Student’s t-test. Source data for panels a, c and d are provided as a Source Data file. Full size image
To further investigate the requirement of extracellular THF for ThfT-mediated SMX resistance and control for any experimental artefacts caused by gene dosage and strain background, we next inserted the thfT T71 variant into the genome of a thfT-negative GAS strain by homologous recombination. For this experiment, we used NS5347 as the parent strain as it belonged to the same multi-locus sequence type (MLST 289) as TB08 and had the same nucleotide sequences flanking the thfT locus. This enabled efficient recombination of thfT into exactly the same locus as in TB08. Examination of each strain in the absence and presence of 50 ng/cm2 exogenous THF revealed that thfT was able to confer high levels of SMX resistance (>1024 μg/ml), but only in the presence of exogenous THF (Fig. 4c, Supplementary Table 5). This resulted in a very modest increase in SXT MIC (~4.5-fold) with no associated increase in TMP MIC. These results demonstrate that ThfT functions as a SMX resistance determinant that imports extracellular THF and bypasses the inhibitory activity of SMX on the folate biosynthesis pathway (Fig. 4b).
If ThfT allows GAS to bypass the folate synthesis pathway altogether in the presence of extracellular THF, then strains that harbor thfT should have complete resistance to SMX in the presence of extracellular THF. As the maximum resolution of the Etest is 1024 μg/ml, we compared relative SMX resistance levels of NS5347 and NS5347::thfT by broth microdilution across a range of SMX concentrations up to 5 mg/ml (the maximum concentration we could dissolve into the growth media) in the absence and presence of THF. Using this method, we found that NS5347::thfT was able to grow in the presence of at least 5 mg/ml of SMX in the presence of exogenous THF but not in the absence of exogenous THF (Fig. 4d). Taken together, these results demonstrate that thfT encodes an antibiotic resistance protein that provides near-absolute resistance to SMX by bypassing the direct inhibitory action of this antibiotic on the folate synthesis pathway.
ThfT enables GAS to acquire multiple one-carbon THF molecules from host cells
In addition to the role of THF as an intermediate in the folate cycle, THF is also used as a carrier for one-carbon molecules in biosynthetic reactions, which are ultimately recycled back to THF7. The differences in the susceptibility of TB08-2-14 to SXT on MHF-Ox and MHF-Bm media suggest that there are differences in the concentration of THF or related compounds in these media. To investigate this, we compared the composition of MH base medium from each supplier using nuclear magnetic resonance (NMR) spectroscopy and reversed phase LC MS/MS approaches (Supplementary Note 1). While we found major differences in the composition of MH base medium from each supplier by both methods, we were unable to identify substantial differences in folate pathway compounds that might explain the relative performance of each media for detecting ThfT-mediated SMX resistance. We next examined the ability of different reduced folate compounds to directly rescue SMX resistance in GAS expressing ThfT. For this experiment, we used the NS5347 strain derivatives encoding the thfT T71 variant inserted as a single gene copy into the chromosome (NS5347::thfT). SMX resistance of each strain was determined by Etest in the presence of 50 ng/cm2 of each different exogenous one-carbon THF compound spread over the surface of the agar plate (Fig. 5a; Supplementary Tables 6 and 7). Expression of ThfT conferred very high SMX resistance to NS5347 in the presence of all one-carbon THF intermediates except for 5-methyl-THF, likely because GAS strains lack homologues of the enzymes required to convert 5-methyl-THF back to THF (Fig. 5b; Supplementary Fig. 8). These results demonstrate that the ThfT T71 variant is able to import multiple one-carbon THF compounds from the extracellular environment.
Fig. 5: ThfT enables SMX-resistance by uptake of multiple one-carbon folate cycle intermediates from host cells. a Ability of NS5347 encoding thfT variants to utilise multiple one carbon THF intermediate compounds. SMX MICs (μg/ml) were determined by Etest in the presence of 50 ng/cm2 of each compound on MHF-Bm agar. Results are median values from three independent experiments. Statistical analysis of groups is in Supplementary Table 7. 5,10-me+THF, 5,10-methenyl-THF. b Model of ThfT mediated SMX resistance in the presence of one-carbon folate cycle intermediate compounds. Compounds utilised by ThfT to bypass the inhibitory action of SMX are highlighted in red and compounds which could not be utilised by ThfT are highlighted in green. 5-formimino-THF (grey) was not available for testing. Solid black arrows indicate pathways that are likely functional in GAS based on the presence of predicted enzymes in the core GAS genome, while pathways indicated with dashed grey arrows indicate the absence of corresponding enzyme43 (Supplementary Fig. 8). Created with BioRender.com. GTP guanosine triphosphate, DHP dihydropteroate, DHF dihydrofolate, dUMP deoxyuridine monophosphate, dTMP deoxythymidine monophosphate, 5,10-me-THF 5,10-methylene-THF. c Cartoon schematic of experiment to test uptake of cellular folates by ThfT. SMX MICs were by broth microdilution across a 2-fold serial dilution of SMX (2500 to 5 μg/ml, left to right) in the presence of a 2-fold serial dilution of epithelial cell lysates (1 × 105 to 1.6 × 103 cells/well, top to bottom). After overnight incubation, MIC values were determined by examining GAS growth (orange wells) for each epithelial cell lysate concentration. d Ability of epithelial cell lysates to confer ThfT-mediated SMX resistance using procedure in c. Data from three biological replicates are presented as mean values ± SEM. Differences assessed using a one-tailed, paired Student’s t-test. Source data for panels a and d are provided as a Source Data file. Full size image
The above results show that ThfT enables uptake of multiple reduced folate compounds that comprise the intracellular folate pool in host cells. We next tested the ability of ThfT to acquire reduced folates directly from primary human epithelial cells. For this experiment, we used a modified broth microdilution assay to allow measurement of SMX MICs in the presence of different concentrations of epithelial cell lysate (Fig. 5c). Results of this experiment show that the cellular folate pool is sufficient to rescue growth of NS5347::thfT but not the NS5347 parental strain (Fig. 5d). Thus, ThfT is able to acquire reduced folates from host cells and bypass SMX inhibition of folate synthesis. We propose that cellular folate cycle intermediates and one-carbon THF pools serve as multiple in vivo substrates for ThfT-mediated SMX resistance (Fig. 5b), which are likely released during infection from neutrophils recruited to the site of infection and/or tissue damage that occurs due to production of one or more GAS cytolysins25,26.
thfT is likely acquired by horizontal transfer from related Streptococcus species
Supplementary Table 1 shows that thfT was variably encoded by the GAS strains examined in this study. To obtain a more comprehensive view on thfT carriage and SXT resistance in GAS strains, we investigated the presence of thfT, dfrF and dfrG in a global dataset of 2083 diverse GAS isolates27 (Supplementary Table 8). Carriage of thfT was uncommon in this dataset (20/2083 GAS strains) yet maintained high levels of sequence homology. Furthermore, thfT-positive strains included 11 unique emm sequence types that were from multiple geographic regions. Of these, 6/20 thfT-positive strains also encoded a TMP resistance gene (dfrF or dfrG; Supplementary Table 8) and are likely functionally SXT resistant in the presence of extracellular reduced folate compounds.
A gene identical to thfT (annotated as folT) was recently described to be carried on a similar genetic element in a subset of GAS, S. dysgalactiae subsp. equisimilis and S. dysgalactiae subsp. dysgalactiae strains28, suggesting a likely common origin. Examination of publicly available S. dysgalactiae genome sequences from refseq revealed that carriage of thfT was also uncommon in this species (3/136 strains; Supplementary Table 9), with no strains identified as carrying both thfT and a TMP resistance gene.